All models are wrong.
Some are useful.
(G. E. P. Box, 1979)
How do we tell which?
What today's math is for
Every time you fit a model to biological data, you face a choice:
- Phylogeny. JC, K80, HKY, GTR, $+\Gamma$, $+I$, … pick the wrong substitution model and your tree is biased.
- Trait evolution. Constant-rate Brownian motion, or an early burst? Or convergence on multiple adaptive peaks? (Next lecture.)
- Divergence times. Strict molecular clock, or a relaxed one? The same calibrations can give different node ages depending on how rate variation is modelled across branches.
A bigger model can always fit the observed data better. The question is:
When does an extra parameter improve a model enough to justify its inclusion?
That is what this lecture answers.
Three tools you'll meet today
- Likelihood Ratio Test. Nested models only; classical $\chi^2$.
- AIC / AICc / AIC weights. Any models; graded support.
- Bayes Factors. Bayesian; integrates over parameter uncertainty.
Next lecture applies all three to evolutionary trait data, and ends with a paper whose punchline is "$\Delta\mathrm{AIC}_c = 162.6$".
Recall: likelihood and the MLE
- From the substitution models lecture (Week 4): a coin tossed 5 times gives $D=(H,T,T,H,T)$.
- The likelihood of the heads probability $f$: $$L(f|D) = f^2(1-f)^3$$
- The maximum likelihood estimate is $\hat{f} = 2/5 = 0.4$ (the observed proportion of heads).
- The same idea applies to any model: find the parameter value that makes the data most probable.
The traditional approach: hypothesis testing
- Classical statistics asks: can we reject a null hypothesis?
- Compute a $p$-value: probability of data as extreme as observed, assuming $H_0$.
- Reject $H_0$ if $p < \alpha$ (conventionally $0.05$).
- For our coin: $H_0$: $f = 0.5$, data $D = (H,T,T,H,T)$.
- Two-sided binomial test: $p = 1.0$.
- We fail to reject $H_0$, and the test has no further answer.
- But the data clearly carries information about $f$: the previous slide gave $\hat{f} = 0.4$.
- And what if we had more data? Would the test tell us what $f$ is, or only whether it differs from $0.5$?
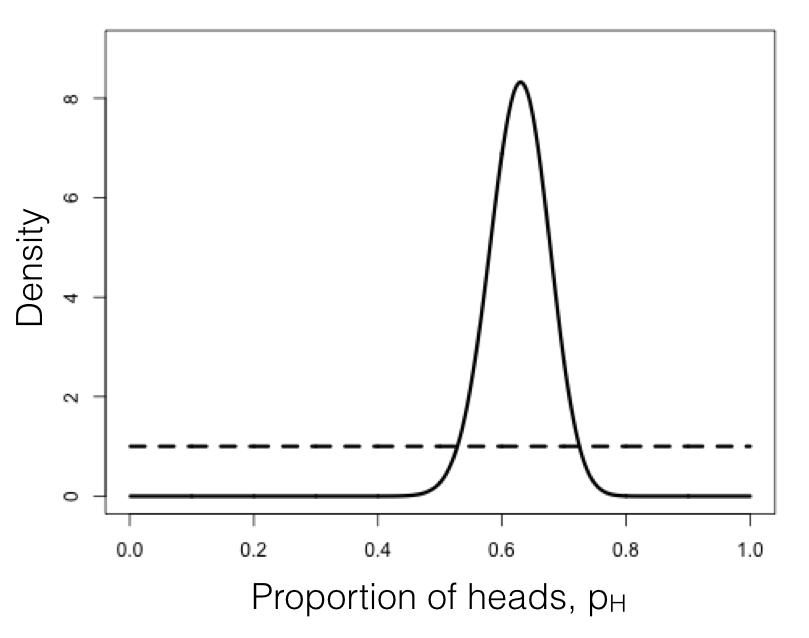
More data: lizard flipping (Harmon, Ch. 2)
- Flip a lizard 100 times: observe 63 heads, 37 tails.
- Two-sided binomial test of $H_0$: $f = 0.5$ gives $p = 0.006$, so we reject: the lizard is unfair.
- But what is $f$? Likelihood: $$L(f|D) = f^{63}(1-f)^{37}$$
- Maximum likelihood estimate: $\hat{f} = 63/100 = 0.63$.
- Same idea as the coin, just with more data the likelihood is much more sharply peaked.
Fitting statistical models to data
- Broadly, parameter estimation is more informative than test-statistics (i.e., $p$-values).
- Rejecting null hypotheses is rarely interesting in biology: the null is often obviously false.
- Recall that the likelihood is: $$L(H|D) = P(D|H)$$
- In an ML framework, the hypothesis with the highest probability of having generated the data is preferred.
- ML will not always return accurate parameter estimates, even when the data is generated under the actual model.
- As with all estimators, ML improves with more data.
Likelihood Ratio Test
- Can only compare two nested models: one must be a special case of the other.
- Test statistic: $\Delta = 2(\ln L_1 - \ln L_0)$, where $L_1$ is the more complex model.
- Under the null, $\Delta \sim \chi^2_k$ where $k$ is the difference in number of parameters.
- Limitations:
- Results can depend on order of tests.
- Cannot compare non-nested models.
Akaike Information Criterion (AIC)
- AIC balances model fit against model complexity: $$\text{AIC} = 2k - 2\ln L$$ where $k$ is the number of estimated parameters.
- AIC assumes that reality is more complex than any of our models: the goal is to identify the model that most efficiently captures the important patterns in the data.
- Lower AIC is better.
- For small sample sizes, use the corrected version: $$\text{AIC}_c = \text{AIC} + \frac{2k(k+1)}{n-k-1}$$ AIC$_c$ is recommended for general use over generic AIC.
AIC weights
- AIC weights provide a more interpretable way to use AIC outputs.
- For model $i$ in a set of $R$ candidate models: $$w_i = \frac{e^{-\Delta_i/2}}{\sum_{r=1}^{R} e^{-\Delta_r/2}}$$ where $\Delta_i = \text{AIC}_i - \text{AIC}_{\min}$.
- Can be interpreted as an estimate of the probability that model $i$ is the best model in the set.
- Modern phylogenetic software frequently calculates AIC for model selection (e.g., ModelFinder in IQ-TREE).
Worked example: choosing a substitution model
A typical alignment fit by maximum likelihood under four nucleotide models. More-parameterised models fit better, but do they fit enough better? Here $k$ counts substitution-model free parameters; branch lengths are separate.
| Model | $k$ | $\ln L$ | AIC | $\Delta$AIC | weight $w_i$ |
|---|---|---|---|---|---|
| JC | 0 | $-2100.0$ | 4200.0 | 33.0 | $\approx 0$ |
| K80 | 1 | $-2085.0$ | 4172.0 | 5.0 | 0.057 |
| HKY | 4 | $-2079.5$ | 4167.0 | 0.0 | 0.690 |
| GTR | 8 | $-2076.5$ | 4169.0 | 2.0 | 0.254 |
This is what IQ-TREE's ModelFinder and jModelTest do automatically, across many more models.
Bayes Factors
- From posterior distributions, models can be compared using Bayes Factors: $$B_{12} = \frac{P(D|H_1)}{P(D|H_2)}$$
- Bayes Factors are ratios of marginal likelihoods $P(D|H)$: the probability of the data averaged over all parameter values, weighted by the prior.
- Unlike ML, this integrates over the full parameter space rather than evaluating only at the best-fit point.
- Tends to favour simpler models than AIC. Assumes the true model is among those tested.
Interpreting Bayes Factors
Kass & Raftery (1995) scale for $2\ln B_{12}$:
| $2\ln B_{12}$ | Evidence for $H_1$ |
|---|---|
| 0 to 2 | Not worth more than a bare mention |
| 2 to 6 | Positive |
| 6 to 10 | Strong |
| > 10 | Very strong |
- For nested models, $2\ln B_{12}$ is on the same scale as the LRT statistic.
- Computing the marginal likelihood is hard. Common methods:
- Path sampling / stepping-stone
- Harmonic mean estimator (unreliable; avoid)
- Nested sampling
Model selection: summary
| Method | Compares | Key assumption |
|---|---|---|
| Likelihood Ratio Test | Nested models only | Null model is true |
| AIC / AIC$_c$ | Any models | Truth is more complex than any model |
| Bayes Factors | Any models | True model is among candidates |
- AIC and Bayes Factors can disagree: Bayes Factors may prefer a simpler model.
- All methods assume good data and correctly specified candidate models.
- Next lecture: we apply these tools to comparative trait data.
Recommended Reading

- Chapter 7: Statistical testing
- Chapter 2: Fitting statistical models